
глюкокортикоїди, прогестерон.
Серед білків, які викликають імунну толерантність організму самки під час вагітності виділяють: α-фетопротеїн, який продукує печінка плода, та α1-глікопротеїн і трофобластичний β2-глікопротеїд, що утворюються у плаценті.
Статевий цикл, роди, післяродовий період супроводжуються змінами епітелію ендометрію (деструкція, епітелізація, проліферація та регенерація), скеровані на обмеження проникнення сперміїв у кров'яне русло та обмеження виходу макрофагів.
Введені в геніталії самки спермії під час осіменіння мають ліпопротеїдну оболонку, оточені плазмою та секретом матки, що обмежує контакт антигену з імунокомпетентними клітинами.
У випадку запліднення відбувається об'єднання генетичного матеріалу самця і самки, формується нова антигенна структура – зигота, яка маскує цей антигенний матеріал прозорою оболонкою. Після розриву прозорої оболонки зростає імуногенність ембріона. Роль імунологічного фільтра тепер виконує трофобласт, що вступає в безпосередній контакт зі слизовою оболонкою матки. Він виділяє супресорний фактор, що захищає ембріон від відторгнення. Розміщені на поверхні трофобласта антигени утворюють комплекси з антитілами, за якими материнський організм розпізнає сумісність ембріона.
Ембріони також продукують гормони, які сигналізують про настання вагітності (ранній фактор вагітності) і проявляють вплив на перебіг ембріогенезу. Саме ці гормони пригнічують синтез у матці простагландинів F2α. Продукований жовтим тілом прогестерон підсилює імуносупресивну дію ендометрію і пригнічує продукцію інтерлейкіну.
До групи причин пов’язаних з розвитком ембріона також відносять і хромосомні аномалії та недостатність фактора ранньої вагітності, після вилуплювання ембріонів. Можливо за розвитку невеликої кількості ембріонів у матці. У результаті цього слизова оболонка матки виділяє в кров простагландин групи F2α, під впливом якого відбувається розсмоктування жовтих тіл у яєчнику (зниження вмісту прогестерону) та як наслідок ембріональна смертність.
Найбільш вирішальним є період, що передує імплантації ембріонів – період судинної реакції матки у місці прикріплення ембріона. Розпочинається на 16–17-й день, тобто коли жовте тіло статевого циклу перетворюється у – вагітності. Судинна реакція матки у цей критичний період вагітності відбувається під впливом неспецифічних факторів ембріонів, яким є естрадіол 17β, що синтезується ембріоном з 13-денного віку. Велике значення мають також імунологічні порушення у системі матки – плацента – плід.
У подальшому імунний захист плода здійснюється змінами в організмі самки і плаценті. Тому вирішальну роль у подальшому збереженні вагітності відіграють плодово-материнські відносини. Плацента і плодові води виконують тепер роль масивного тканинного бар’єра між плодом і материнським організмом.
Амніотична та алантоїсна рідини відзначаються антигенною подібністю з ембріоном, тому вони зв'язують скеровані проти нього антитіла. З появою плаценти встановлюється імунна рівновага між ембріоном (плодом) та материнським організмом.
Під час вагітності зростає продукція Т-лімфоцитів (супресорів), що пригнічують синтез В-клітинами антитіл, збільшується в крові вміст кортикостероїдів (інгібують синтез антитіл) і встановлюється рівновага між факторами стимулювання та пригнічення синтезу антитіл.
З наближенням родів порушується імунологічна рівновага і замість толерантності розвиваються реакції відторгнення. У самок усі аномалії яйцеводів, матки, шийки матки і піхви об’єднуються в групу порушень розвитку мюлерових каналів.
Серед генетичних аномалій найбільш поширеними є:
- крипторхізм;
- гіпоплазія гонад (яєчників і сім’яників);
- гермафродитизм;
- подвоєння шийки матки;
- антифемінічні та антимаскулінічні летальні фактори;
- порушення плодючості внаслідок утворення мозаїків і химер.
У самців виділяють декілька груп генетичних аномалій геніталій:
- “Справжні” кісти придатків сім’яників, включаючи спермостаз.
- Парадидиміс (залишки канальців проміжної нирки).
- Мюлерові кісти (Uterovagina masculina – чоловіча матка і чоловіча піхва).
- Кісти всередині і навколо міхурцеподібних залоз.
- Кісти передміхурової і куперової залоз.
Генетичні причини доводиться припускати в тому випадку, коли відповідні порушення проявляються при бездоганному догляді, утриманні, годівлі та за відсутності перевантаження і хвороб. В якості порушень плодючості, що спостерігають у самців, відмічають ще різноманітні генетично обумовлені дефекти сперміїв.
Детальна класифікація аномалій сперміїв зображена на рисунку 1.
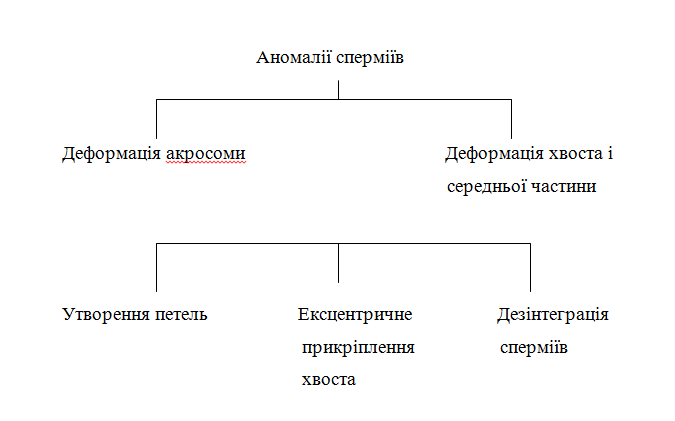
Рисунок 1 – Класифікація аномалій сперміїв
Описані дефекти акросоми сперміїв у вигляді вакуолізації, однак без змін у наборі хромосом.
До групи дефектів хвоста і середньої частини відноситься виявлений на сперміях так званий Даг-ефект – утворення вузьких петель і складок з розщепленням і відсутністю волокон. При ексцентричному прикріпленні хвоста (corkserew tail) спостерігається також його штопороподібне закручування.
Припускається генетична обумовленість. При дезінтеграції сперміїв йдеться про відділення головок з потовщенням середньої частини. Під час проходження через дистальні відділи придатка сім’яника відбувається відділення головок. В середній частині знаходиться крапля цитоплазми, яка переміщується.
До цієї групи дефектів також відноситься ізольоване потовщення середньої частини (“pseudo-droplet”). Припускається, що ці аномалії зумовлені простими автосомними рецесивними, обмеженими статтю факторами.
Крім того, існує велика кількість повідомлень про генетичну обумовленість неплідності тварин. Так, у США спостерігали неплідність самок зумовлену відсутністю охоти (алібідний статевий цикл).
Враховуючи значущість вищенаведеного матеріалу не слід забувати про те, що свиноматки та кнурі за будь-яких обставин мають бути забезпечені повноцінним раціоном, лише за цієї обставини ми можемо вести мову про отримання максимального результату та впливати на зміни в організмі тварин.
БАБАНЬ О.А., канд. вет. наук, консультант чеської компанії “ТЕКРО“
